| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| ЛИМФОЦИТЫ И ИММУНОКОМПЕТЕНТНАЯ СИСТЕМА
Содержание Список сокращений……………………………………………………………4 ЛИМФОЦИТЫ И ИММУНОКОМПЕТЕНТНАЯ СИСТЕМА (морфология и физиология)…………………………………………………. 5 ОЦЕНКА БОЛЬНОГО С ЛИМФАДЕНОПАТИЕЙ……………………...28 ИНФЕКЦИОННЫЙ МОНОНУКЛЕОЗ……………………………………38 ГЕМАТОЛОГИЧЕСКИЕ ЗАБОЛЕВАНИЯ, СОПРОВОЖДАЮЩИЕСЯ УВЕЛИЧЕНИЕМ ЛИМФОУЗЛОВ…………………………………………54 Болезнь Ходжкина……………………………………………………………..54 Неходжкинские лимфомы …………………………………………………….66 Хронический лимфолейкоз………………………………………………………...78 САРКОИДОЗ…………………………………………………………………..89 ГИСТИОЦИТОЗЫ…………………………………………………………..105 Приложение №1. Иммуноклассификация лимфом ……………………108 Приложение №2. Хромосомные нарушения при лимфомах…………..113 Приложение №3. Примеры формулировки диагноза…………………..119 Приложение №4. Ситуационные задачи …………………………………120 ЛИТЕРАТУРА………………………………………………………………..124
Список сокращений CD – кластер дифференцировки FISH – флуоресцентная in situ гибридизация HLA – человеческие лейкоцитарные антигены (главный комплекс гистосовместимости человека) Ig – иммуноглобулин IL – интерлейкин INF – интерферон MALT – лимфоидная ткань слизистых TCR – Т-клеточный рецептор TNF – фактор некроза опухоли АПК – антигенпрезентирующие клетки БАЛ – бронхоальвеолярный лаваж ВЭБ – вирус Эпштейна-Барра ГЗТ – гиперчувствительность замедленного типа ЛГМ – лимфогранулематоз (лимфома Ходжкина) ЛПС – липополисахариды НХЛ – неходжкинские лимфомы ПАЛО – периартериальная лимфатическая оболочка ПЦР – полимеразная цепная реакция РКТ – рентгеновская компьютерная томография высокого разрешения ХЛЛ–хронический лимфолейкоз ЭКВС – эндотелиальные клетки венозных синусов
ЛИМФОЦИТЫ И ИММУНОКОМПЕТЕНТНАЯ СИСТЕМА (морфология и физиология) Иммунокомпетентная система, морфологическим субстратом которой является лимфатическая ткань, поддерживает гомеостаз организма путем специфического распознавания и обезвреживания чужеродных субстанций. Специфическое распознавание чужеродных веществ клетками иммунной системы приводит к активации этих клеток и развитию иммунного ответа. Различают два основных типа иммунного ответа: - гуморальный (чужеродные вещества обезвреживаются антителами; антитела и комплемент убивают клетки и вызывают воспаление); - клеточный (чужеродные или измененные собственные клетки убиваются лимфоцитами). Иммунокомпетентная система представляет собой наиболее зрелое (дифференцированное) звено иммунной системы, способное к осуществлению различного рода иммунологических реакций. В составе иммунной системы есть и «иммунологически не компетентное» звено, которое включает центральные органы иммуногенеза – костный мозг и тимус (рис. 1). Лимфоидные клетки костного мозга и тимуса представлены, в основном, незрелыми и дифференцирующимися предшественниками иммуноцитов (иммунокомпетентных клеток), в большинстве своем не способными осуществлять иммунологические реакции. К числу периферических органов иммунной системы относятся инкапсулированные органы (селезенка, лимфатические узлы), неинкапсулированная лимфоидная ткань, связанная со слизистыми оболочками, кожей и другими органами, а также диссеминированный пул зрелых лимфоидных клеток крови и лимфы. Главной клеткой иммунной системы является лимфоцит. Выделяют популяции лимфоцитов: - Т-лимфоциты, - В-лимфоциты, - К-клетки, - NK-клетки. ЛИМФА
Рис. 1. Органы иммунной системы (схема). Т-лимфоциты участвуют, преимущественно, в клеточном иммунитете и регулируют уровень гуморального иммунитета. В-лимфоциты и их наиболее зрелые формы – плазматические клетки – продуцируют иммуноглобулины (антитела), то есть осуществляют синтез эффекторов гуморального иммунитета. К-клетки не имеют специфических рецепторов для антигена и осуществляют свое действие при посредстве антител. NК-клетки способны специфически распознавать клетки-мишени и лизировать их без участия антител. Т- и В-лимфоциты для осуществления своих функций должны распознать антиген и перейти в активное состояние. Этот процесс осуществляется при помощи Т- и В-клеточных рецепторных комплексов, присутствующих на мембране лимфоцитов (рис. 2). В состав этих комплексов наряду со специфическими рецепторами антигена входят так называемые трансдьюсерные молекулы, то есть молекулы, обеспечивающие передачу сигнала внутрь клетки.
2 – интегральные белки; 3 – полуинтегральные белки; 4 – поверхностные белки; 5 – гликокаликс.
Специфическими рецепторами В-лимфоцитов являются молекулы иммуноглобулинов. На В-лимфоцитах Ig-рецепторы нековалентно ассоциированы с двумя трансмембранными белками - Igα и Igβ или Igα и Igγ. Молекулы Ig и полипептидные цепи, входящие в состав В-клеточного рецептора, являются наиболее надежными маркерами В-линейной принадлежности. Т-клетки, так же, как и В-клетки, способны специфически распознавать антиген. Опознание антигена производится Т-клеточным рецептором, который по своей структуре во многом напоминает молекулу Ig, а точнее - Fab-фрагмент Ig. Т-клеточные рецепторы могут существовать в двух гетеродимерных формах – αβ (большинство периферических Т-клеток) и γδ (примерно 5% периферических Т-клеток). Каждая цепь состоит из 2 доменов – один «константный», другой – «вариабельный». Только при объединении обеих цепей рецептор приобретает функциональную специфичность. Передача сигнала, ведущего к активации Т-клеток после связывания TCR (Т-клеточный рецептор) с презентируемым антигеном, осуществляется комплексом CD3. В составе CD3 присутствуют 3 полипептидных цепи: γ, δ, ε. TCR, CD3 и сигнальные двухцепочечные молекулы (ζζ) образуют Т-клеточный рецепторный комплекс. Для активации Т- и В-лимфоцитов необходимы межклеточные контакты, а также растворимые факторы, называемые цитокинами. В ходе межклеточных контактов происходит распознавание антигена и получение Т-клеткой так называемых костимуляторных сигналов. Цитокины – это продуцируемые клетками небольшие белковые молекулы, активно участвующие в межклеточных взаимодействиях и неспецифичные к антигену. Это колониестимулирующие факторы, интерлейкины, интерфероны и факторы некроза опухоли. В настоящее время известно более 20-ти различных цитокинов. Межклеточные взаимодействия в ходе иммунного ответа играют решающую роль в процессах распознавания чужеродных антигенов, активации и дифференцировки лимфоидных клеток. Т-клетка распознает антиген в комплексе с молекулами HLA. Функция молекул HLA состоит в том, чтобы связывать пептидные фрагменты, происходящие из патогенов, и перемещать их на клеточную мембрану для распознавания соответствующими Т-клетками. Последствия распознавания Т-клетками комплекса пептид-HLA практически всегда губительны для патогена: убийство инфицированных вирусом клеток, активация микробицидного действия макрофагов в отношении внутриклеточных бактерий, активация В-клеток для продукции антител, способных элиминировать или нейтрализовать внеклеточные патогены. Т-хелперные лимфоциты имеют на мембране рецептор CD4 для молекул HLA-II, а Т-цитотоксические лимфоциты имеют рецептор CD8 для молекул HLA-I. Другая популяция Т-хелперов (Тн2) усиливает специфические иммунные ответы, поддерживая пролиферацию и дифференцировку В-лимфоцитов. Помимо киллерного действия, активации макрофагов и В-клеток, Т-лимфоциты (Т-супрессоры) способны специфически подавлять активность других Т-клеток, то есть снижать силу иммунного ответа или индуцировать толерантность. В-лимфоциты также способны перерабатывать антиген, поступивший в клетку посредством иммуноглобулиновых рецепторов, и презентировать его Т-хелперам (Тн2). Лимфоциты в костном мозге В костном мозге человека после рождения происходят наиболее ранние этапы дифференцировки Т- и В-лимфоцитов. Незрелые предшественники Т- и В-лимфоцитов (протимоциты, про-В-лимфоциты, пре-пре-В-лимфоциты, пре-В-лимфоциты) образуются только в костном мозге. Для развития в зрелые Т-клетки или В-клетки примитивные костномозговые клетки-предшественницы нуждаются в специфическом микроокружении. Необходимыми являются межклеточные контакты (в тимусе, костном мозге) и продукция специфических цитокинов: IL-7(интерлейкин-7) для раннего развития В-клеток, IL-2 (интерлейкин-2) для Т-клеток. Костномозговой В-лимфопоэз. В-лимфоцитопоэз характеризуется последовательными событиями, заключающимися в формировании специфических рецепторов (Ig), избирательной экспрессией мембранных и цитоплазматических маркеров, изменением чувствительности к ростовым и дифференцировочным сигналам, выделяемым стромальными клетками и цитокинам. Созревание В-клеточного рецептора. Генерация антител во многом обусловлена соматическими комбинаторными механизмами на уровне генных сегментов ДНК. Молекула иммуноглобулина состоит из 2 легких и 2 тяжелых полипептидных цепей. Известны 2 типа легких (L) цепей (к и λ) и 5 классов тяжелых (Н) цепей (μ, γ, α, δ, ε), а также их подтипы и подклассы. В молекуле Ig имеется 2 идентичные L-цепи и 2 идентичные Н-цепи. Строение легких и тяжелых цепей Ig различное. Тип или класс иммуноглобулина обусловлен константной (С) областью Н- и L-цепей, а его специфичность - вариабельной (V) областью. При объединении вариабельных областей Н- и L-цепей в молекуле Ig образуется 2 специфических антигенсвязывающих участка. Последовательность появления В-клеточных антигенов и рецепторов на мембране костномозговых В-лимфоцитов такова: CD19 -> CD24 -> CD10 -> CD20 -> CD22 -> clg(μ)-> CD21-> sIgM -> slg(M+D). Процесс дифференцировки В-клеток в костном мозге регулируется стромальными клетками костного мозга посредством адгезионных взаимодействий и продукции цитокинов. Важную роль в поддержании пролиферации и дифференцировки ранних предшественников В-клеток играют растворимые факторы и цитокины (IL-7, c-kit, IL-11, IGF), которые продуцируются клетками стромы. Конечные этапы созревания В-лимфоцитов в костном мозге соответствуют незрелым В-клеткам (мембранные IgM) и зрелым девственным (virgin) В-клеткам (мембранные IgM+lgD). В-клетка покидает костный мозг, имея сформированный рецепторный иммуноглобулиновый аппарат (коэкспрессия мембранных IgM и IgD), причем активационные антигены CD23 и CD5 на мембране отсутствуют. Костномозговой Т-лимфопоэз. В организме взрослого человека продукция предшественника Т-клеток происходит в костном мозге. И лишь затем они заселяют тимус и далее – периферические лимфоидные ткани. Плюрипотентные клетки-предшественницы, способные давать начало всем росткам кроветворения, сосредоточены в костном мозге во фракции лимфоидных клеток, экспрессирующих на мембране антиген стволовых гемопоэтических клеток (CD34). Эти клетки способны давать начало также стромальным клеткам. Появление на мембране CD34+ клеток HLA-DR антигенов свидетельствует об их направленности к гемопоэтической дифференцировке. Лимфоциты и тимус Незрелые предшественники Т-лимфоцитов из костного мозга с током крови поступают в тимус. Становление репертуара Т-лимфоцитов происходит в тимусе. В тимусе формируется набор специфичностей Т-лимфоцитов, участвующих в эффекторных реакциях в периферических лимфоидных тканях. Тимус (вилочковая железа) является лимфоидно-эпителиальным органом. Основным структурным элементом тимуса является тимическая долька (рис. 3). Процессы внутритимической дифференцировки и селекции происходят в различных отделах дольки. В тимусе представлен широкий спектр лимфоидных и эпителиальных клеток: субкапсулярные тимические бласты, кортикальные тимоциты, медуллярные тимоциты, тельца Гассаля, дендритные клетки и т.д.
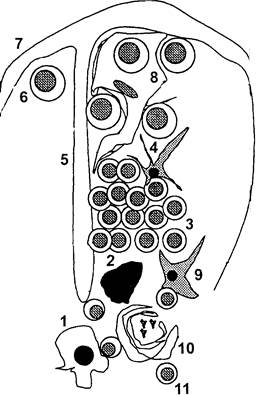
Рис. 3. Структура тимической дольки. 1 – медуллярные эпителиальные клетки; 2 – макрофаг; 3 – агрегаты кортикальных тимоцитов; 4 – кортикальная дендритная клетка; 5 – септа; 6 – лимфобласт; 7 – капсула; 8 – клетка-нянька; 9 – интердигитирующая клетка; 10 – тельце Гассаля; 11 – медуллярный тимоцит.
Тимоцитарная пролиферация предшествует экспрессии TCRαβ. При поступлении в тимус протимоциты становятся коммитированными по Т-клеточной линии и начинают экспрессировать CD3 в цитоплазме (cCD3) и TdT. Эти незрелые тимоциты являются крупными бластами и клетками среднего размера с высокими уровнями экспрессии TdT и самой большой пролиферативной активностью среди всех тимоцитов. На следующем этапе дифференцировки обычные (common) или кортикальные тимоциты являются cCD3+ TdT-. Они имеют средний уровень пролиферативной активности и отличаются по появлению цитоплазматической экспрессии TCRβ. В кортикальных тимоцитах только начинается перестройка α-генов и TCRαβ еще не экспрессирован. Клетки имеют на мембране CD4 и CD8, CD1+, CD2+. На поздних стадиях тимического развития зрелые (медуллярные) тимоциты экспрессируют либо CD4 либо CD8, мембранные CD3+ (mCD3+), CD2+, TCRαβ+; TdT и CD1 утрачиваются. Таким образом, тимус является центральным органом иммунной системы, ответственным за образование пула наивных Т-лимфоцитов. Позитивно отобранные клетки в итоге покидают тимус и патрулируют тело в качестве длительно живущих покоящихся лимфоцитов. Их реактивация может произойти, как правило, только под влиянием чужеродных пептидов, представленных собственными молекулами HLA. |



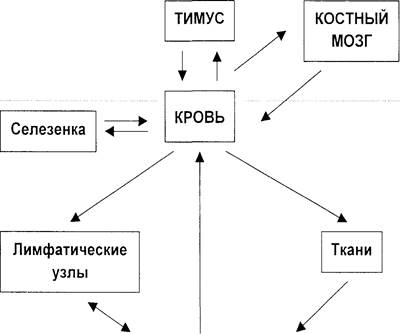
 Рис. 2. Модель строения плазматической мембраны (по Singer, Nicolson, 1972; рис. И.Н. Першиной). 1 – липидный слой;
Рис. 2. Модель строения плазматической мембраны (по Singer, Nicolson, 1972; рис. И.Н. Першиной). 1 – липидный слой;