| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Реципрокная рекомбинация и конверсия
В ходе синапсиса гомологичные хромосомы обмениваются фрагментами. В световом микроскопе последствия этого обмена видны как перекресты, или хиазмы.
Рисунок 22. Хиазмы в результате трех отдельных перекрестов хроматид обеих хромосом и последующее расхождение хромосом Частота рекомбинаций определяется по формуле Частота рекомбинаций генов показывает относительное расположение сцепленных генов в хромосоме: чем дальше друг от друга находятся гены, тем выше частота рекомбинации. Это обстоятельство используется при составлении генетических карт. Условное «расстояние» между локусами двух генов считается прямо пропорциональным частоте рекомбинации. Взаимное расположение локусов трёх и более генов определяется методом триангуляции. При этом сначала берутся гены с наименьшей частотой рекомбинации. Далее выбирают следующую по величине частоту рекомбинации и указывают два возможных положения нового гена; одно из этих положений будет отсеяно на следующем шаге, когда берётся третья частота. В реальных экспериментах генетические карты могут искажаться благодаря двойному кроссинговеру, когда рекомбинация происходит одновременно в двух точках. Двойной кроссинговер особенно характерен для генов, локусы которых разделены большими расстояниями В настоящее время общепризнаны две модели рекомбинации, приводящие к конверсии. Одна из них - модель Р. Холлидея, вторая - Дж. Жостака и др., в которой рекомбинация начинается с двуцепочечного разрыва в одном из гомологов. Для краткости изложения ограничимся демонстрацией конверсии на примере модели Холлидея. В рисунок 23 включены только те ее этапы, которые непосредственно относятся к конверсии. Если в тетраде произошел кроссинговер в участке гена В, то соотношение аллелей генов А и С, внешних (фланговых) по отношению к участку кроссинговера, не изменяется и остается равным 2А : 2а и 2С : 2с, то есть рекомбинация по ним реципрокна. Напротив, рекомбинация маркеров, попавших непосредственно в участок кроссинговера, то есть в гетеродуплекс, обычно нереципрокна из-за коррекции неспаренных оснований, внесенных разными аллелями (B и b). Протяженность гетеродуплекса в мейотической рекомбинации достигает 1 т.п.н., в случае митотической рекомбинации она еще больше. Все гетерозиготные маркеры, попавшие в гетеродуплекс, обычно конвертируют совместно, что является следствием существования участка конверсии. Этот факт основан на работе рассмотренной выше системы коррекции с вырезанием и застройкой длинных брешей.
А теперь выделим другие основные закономерности процесса конверсии. Конверсия обычно равнонаправленна. Выбор удаляемого основания из двух неспаренных случаен, поэтому тетрады 3B : 1b и 1B : 3b равновероятны. Конверсия полярна. Это проявляется в том, что частоты конверсии разных аллелей одного гена закономерно снижаются, начиная с определенной точки, чаще всего от одного конца гена к другому. У дрожжей они обычно варьируют от 18% до долей процента. Так как частота конверсии аллеля зависит от вероятности его попадания в гетеродуплекс, то полярность указывает на наличие в хромосомах фиксированных сайтов, в которых инициируется рекомбинация. В последние годы у дрожжей идентифицировано несколько таких сайтов. Их называют горячими точками рекомбинации.
Проектное задание 1. Опишите расположение хромосом в метафазной пластинке в клетке с 2n=6 во время (а) митоза, (б) мейоза I и (с) мейоза II. 2. Диплоидная клетка содержит три пары гомологичных хромосом: С1 С2, М1 М2 и Е1 Е2. Каковы возможные комбинации этих хромосом в отсутствие кроссинговера (а) в двух дочерних клетках после митоза, (б) в метафазе I мейоза, (с) в гаплоидных клетках после делений мейоза? 3. В результате мутации клетка А и клетка Б потеряли способность синтезировать ДНК-полимеразу. Какова вероятность передачи этой мутации дочерним клеткам, если она произошла у клетки А в период G1, а у клетки Б – в период G2 митотического цикла? 4. В клетках А и Б в интерфазе возник мутантный ген. Они нормально завершили митотический цикл, но после митоза клетки А обе дочерние клетки получили мутантный ген, а после митоза клетки Б мутантный ген оказался в одной из дочерних клеток. Чем это можно объяснить? 5. Гены, которые должны были включиться в работу в периоде G2, остались неактивными. Отразится ли это на ходе митоза? 6. В результате действия колхицина в течение двух митотических делений получены клетки ржи с 56 хромосомами. Каково гаплоидное число хромосом у ржи? 7. Почему гаплоидные растения томатов обычно не дают семян и размножаются только вегетативно? В каком исключительном случае гаплоидный томат может дать семена? 8. Если соматическая клетка имеет 28 хромосом, то сколько хроматид идет к каждому полюсу в анафазе эквационного деления? Каково число бивалентов в профазе редукционного деления? 9. Скрестили тетраплоидный вид пшеницы (28 хромосом) с тетраплоидным видом эгилопса (28 хромосом) (дикий родственник пшеницы). Полученные гибридные семена высеяли и получили гибридов первого поколения. В мейозе у данных гибридов наблюдали образование 7 бивалентов, остальные хромосомы были в виде унивалентов. Что вы можете сказать о хромосомах данных видов, о степени их родства? 10. Какое расположение спор в аске нейроспоры доказывает, что кроссинговер осуществляется на стадии 4 хроматид? Объясните. ААААаааа; ааааАААА; ААааааАА; ааААааАА
Тесты рубежного контроля 1. В синтетический период интерфазы происходит:а) удвоение пластид и митохондрий; б) синтез ДНК и р-РНК; в) синтез АТФ и белков; г) накопление нуклеотидов ДНК, синтез и-РНК и белков; д) синтез белков ахроматинового веретена и ДНК. 2. В постсинтетический период интерфазы происходит:а) синтез ДНК и ферментов; б) синтез ДНК, р-РНК, рост клетки; в) синтез АТФ; г) накопление нуклеотидов ДНК; д) синтез белков ахроматинового веретена. 3.Содержание генетического материала в клетке в пресинтетический период интерфазы:а) 1n1chr1с; б) 1n2chr2с; в) 2n1chr2с; г) 2n2chr4с; д) 1nbiv4chr4с. 4. Основные причины митоза:а) увеличение ядерно-цитоплазматического отношения; б) уменьшение ядерно-цитоплазматического отношения; в) репликация молекулы ДНК и «раневые гормоны»; г) "раневые гормоны" и митогенетические лучи; д) нарушение целостности кариолеммы. 5. Мейозом делятся клетки:а) соматические и стареющие; б) половые и клетки эмбриона; в) гаметоциты; г) клетки опухолей; д) клетки регенерирующих тканей. 6. В профазу мейоза I происходит:а) спирализация хроматина; б) деспирализация хромосом; в) удвоение центросом; г) конъюгация хромосом; д) кроссинговер. 7. В анафазу мейоза I происходит:а) спирализация хроматина; б) деспирализация хромосом; в) расхождение гомологичных хромосом к полюсам; г) конъюгация хромосом; д) кроссинговер. 8.К митотическому аппарату клетки относятся: А. Хромосомы, центриоли, нити веретена деления Б. Митохондрии, рибосомы, микротрубочки Г. Хромосомы, комплекс Гольджи, лизосомы 9.Движение хромосом к полюсам клетки осуществляется за счет: А. Циклоза Б. Сокращения хромосом В. Сокращения нитей веретена деления 10. На какой стадии первого деления мейоза происходит образование бивалентов и кроссинговер: А. Профаза, метафаза Б. Лептотена, зиготена, В. Зиготена, пахитена, Г. Пахитена, диплотена |





 , где N – количество рекомбинантов,
, где N – количество рекомбинантов, 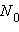 – общее количество потомков. В то же время частота рекомбинаций определяет число рекомбинаций, происходящих при образовании гамет.
– общее количество потомков. В то же время частота рекомбинаций определяет число рекомбинаций, происходящих при образовании гамет.