| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Хемомеханический этап мышечного сокращения.
Теория хемомеханического этапа мышечного сокращения была разработана О. Хаксли в 1954 г. и дополнена в 1963 г. М. Девисом. Основные положения этой теории: 1) ионы Ca запускают механизм мышечного сокращения; 2) за счет ионов Ca происходит скольжение тонких актиновых нитей по отношению к миозиновым. В покое, когда ионов Ca мало, скольжения не происходит, потому что этому препятствуют молекулы тропонина и отрицательно заряды АТФ, АТФ-азы и АДФ. Повышенная концентрация ионов Ca происходит за счет поступления его из межфибриллярного пространства. При этом происходит ряд реакций с участием ионов Ca: 1) Ca2+ реагирует с трипонином; 2) Ca2+ активирует АТФ-азу; 3) Ca2+ снимает заряды с АДФ, АТФ, АТФ-азы. Взаимодействие ионов Ca с тропонином приводит к изменению расположения последнего на актиновой нити, открываются активные центры тонкой протофибриллы. За счет них формируются поперечные мостики между актином и миозином, которые перемещают актиновую нить в промежутки между миозиновой нитью. При перемещении актиновой нити относительно миозиновой происходит сокращение мышечной ткани. Итак, главную роль в механизме мышечного сокращения играют белок тропонин, который закрывает активные центры тонкой протофибриллы и ионы Ca.
№21 Одиночное мышечное сокращение, суммация одиночных мышечных сокращений. Одиночное мышечное сокращение – сокращение мышцы в овеет на одиночный стимул. График Одиночного мышечного сокращения: 3 фазы: I скрытый латентный период. Распространение потенциала действия, заход в трубочки; II фаза сокращения, укорочения, напряжения; III нисходящая фаза – расслабления.
Суммация одиночного мышечного сокращения, если повторный стимул попадает в фазу сокращения – полная суммация, амплитуда будет выше и только одна вершина; в фазу расслабления – неполная суммация, 2 отдельно стоящие вершины.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверх порогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
№22 Понятие о тетанусе, виды и механизмы тетануса. Оптимум – тетанус максимальной амплитуды. Пессимум частоты – такая частота, при которой мышца перестает отвечать на раздражение и расслабляется. Непрямое раздражение опосредуется через нерв. Прямое при условии: - если электрод введен непосредственно в мышцу; - или же блокируем нервные окончания в мышце (ядом Курара).
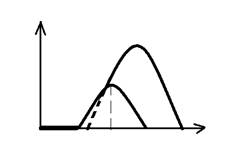
Тетанус- это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация (рис).. Гладкий тетанус возникает тогда, когда', каждое последующее раздражение наносится а конце периода укорочения т.е. имеет место полная суммация отдельных сокращений и (рис.). Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например тремор рук; при алкогольной интоксикации и болезни Паркинсона.
Различают два вида тетанических сокращений или тетануса: зубчатый и гладкий (сплошной). Зубчатый тетанус наблюдается тогда, когда последующий импульс приходит в фазу расслабления мышцы (состояние мышцы чисто лабораторное). Гладкий тетанус имеет место тогда, когда очередной импульс попадает в конце фазы укорочения. Величина тетанического сокращения, как и одиночного, подчиняется закону градуальности или силовых отношений: чем больше сила раздражителя и частота импульсов, тем сильнее сокращается мышца. Механизмы увеличения тетанического сокращения точно такие же, как и для одиночного, то есть зависят от вида нейромоторных единиц (суммация в пределах нескольких или одного мышечного волокон). Известным отечественным физиологом Н. Е. Введенским были выявлены определенные закономерности силы мышечного сокращения, связанные, с одной стороны, с силой раздражителя, с другой, с частотой. В связи с этим Введенский вводит в физиологию такие понятия как пессимум и оптимум частоты и силы раздражителя. Так, под оптимумом силы раздражителя понимают такой по силе раздражитель, при действии которого на мышечную ткань возникает максимальный сократительный эффект. Под пессимумом раздражителя понимают такой по силе раздражитель, который вместо увеличения силы мышечного сокращения вызывает его уменьшение. Например, подается раздражитель силой 10 В, который вызывает максимальный сократительный эффект, а увеличение силы этого параметра до 15 В сопровождается уменьшением силы сокращения. Однако, следует помнить, что в целостном организме сила раздражителя практически остается одна и та же, т. к. амплитуда потенциала действия практически не меняется (100-120 мВ). Поэтому с точки зрения физиологии следует говорить не об оптимуме и пессимуме силы раздражителя, а об оптимуме и пессимуме частоты раздражителя. Под оптимумом частоты раздражителя следует понимать ту наибольшую частоту раздражителя, при действии которой на мышцу наблюдается ее максимальное сокращение. Под пессимумом частоты раздражителя подразумевают такую частоту раздражителя, при действии которой на мышцу вместо увеличения сократительного эффекта имеет место его уменьшение. Например, частота раздражителя 150 им/с вызывает максимальное сокращение мышцы, увеличение же частоты посылаемых сигналов до 200 им/с сопровождается уменьшением сократительного эффекта мышцы. Максимальное сокращение мышцы наблюдается тогда, когда частота поступающих к ним импульсов соответствует лабильности ткани. Если же частота адресующихся к мышце сигналов превосходит лабильность ткани, то развивается пессимум частоты, т. к. часть импульсов попадает в период рефрактерности и не проявляет своего действия на эффектор. Принято считать, тетаническое сокращение является суммой одиночных сокращений, которые возникают в ответ на каждый импульс. Однако, исследования показали, что сумма площади тетанического сокращения больше, чем арифметическая сумма одиночных сокращений. Это значит, что в определенные моменты тетанического сокращения последующий сократительный эффект больше предыдущего. Запись тетанического сокращения действительно показала, что вначале тетанус несколько увеличивается, а не носит характера горизонтального плато. Происходит, так называемое, суперпозиция - наложение одного сокращения на другое, имеющее место только в начале тетанического сокращения. Для объяснения суперпозиции в физиологии имеется три точки зрения. 1. Теория, предложенная Введенским. Сущность теории: суперпозиция может быть лишь в том случае, если последующий импульс попадает в фазу повышенной возбудимости (экзальтации). Действительно, в фазу повышенной возбудимости каждый пороговый раздражитель превращается в надпороговый, поэтому и вызывает эффект мышечного сокращения больше порогового. 2. Теория, предложенная Бабским. Согласно этой теории суперпозиция связана с изменением метаболизма в мышце. Бабский полагал, то при действии на мышцу первого импульса образуется очень много источников энергии в виде АТФ, которые не успевает расходоваться при первом возбуждении и они используются в последующем возбуждении, приводя к увеличению силы мышечного сокращения. 3. Теория, связывающая суперпозицию с ионами Са. Согласно этой теории при поступлении первого импульса выделяется избыточное количество ионов Са из саркоплазматического ретикулюма, который не успевает весь использоваться в сократительном акте. Его избыточное количество приводит к увеличению силы последующих сокращений. Тонические сокращения являются фактически разновидностью тетанических сокращений, представляющие собой длительное укорочение мышц и обусловливающие в основном мышечный тонус - постоянное незначительное напряжение мышц, имеющий место в мышечной ткани в состоянии покоя. Это постоянное напряжение мышечной ткани имеет место даже в состоянии сна. Если в эксперименте произвести денервацию мышц, то тонус их исчезает, что указывает на его нервную природу. Кроме тетанических и тонических сокращений мышц различают обратимые и необратимые контрактуры.Контрактура - это длительное укорочение мышечной ткани, которое имеет место, например, при утомлении и носит обратимый характер. Необратимые контрактуры наблюдаются при трупном окоченении или действии высоких температур. В последнем случае имеет место денатурация сократительного белка. Известно, что после наступления смерти наблюдается трупное окоченение - длительное напряжение мышц, связанное с тем, что исчезает АТФ, необходимая для возвращения ионов Са в саркоплазматический ретикулюм.
№ 23 Типы мышечных сокращений. Различают такие виды сокращения мышц как одиночные, тетанические и тонические. Различают следующие режимы мышечного сокращения: Различают два типа мышечных сокращений. Если оба конца мышцы неподвижно закреплены, происходит изометрическое сокращение, и при неизменной длине напряжение увеличивается. Если один конец мышцы свободен, то в процессе сокращения длина мышцы уменьшится, а напряжение не изменяется — такое сокращение называют изотоническим; в организме такие сокращения имеют большее значение для выполнения любых движений. В результате сокращения в мышечных волокнах возникает напряжение. Эта способность развивать напряжение является основным свойством мышцы. Напряжение, развиваемое мышцами при сокращении, реализуется по-разному. Если внешняя нагрузка меньше, чем напряжение сокращающейся мышцы, то мышца укорачивается и вызывает движение – это изотонический (концентрический, миометрический) тип сокращения. Поскольку в экспериментальных условиях при электрическом раздражении укорочение мышцы происходит при постоянном напряжении = внешней нагрузке. Если внешняя нагрузка больше, чем напряжение, развиваемое мышцей во время сокращения, то такая мышца растягивается при сокращении – это эксцентрический (плиометрический) тип сокращения. Изотонический и эксцентрический типы сокращения, т.е. сокращения. при которых мышца изменяет свою длину, относятся к динамической форме сокращения. Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим. Это статическая форма сокращения. Она возникает в 2-ух случаях: либо когда внешняя нагрузка = напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки. При динамических формах сокращения производится внешняя работа – при изотоническом сокращении положительная, при эксцентрическом отрицательная. Величина работы в обоих случаях может быть определена как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" = 0, и, согласно физическому закону, в этом случае мышца не производит никакой работы. Если внешняя нагрузка на мышцы меньше, чем ее напряжение, мышца укорачивается и вызывает движение. Это концентрический, или миометрический, тип сокращения. В экспериментальных условиях при электрическом раздражении изолированной мышцы ее укорочение происходит при постоянном напряжении, равном внешней нагрузке. Поэтому этот тип сокращения называют также изотоническим (изос - равный, тонус - напряжение). Если внешняя нагрузка на мышцы больше, чем ее напряжение, развиваемое во время сокращения, мышца растягивается (удлиняется). Это эксцентрический, или плиометрический, тип сокращения. Концентрический и эксцентрический типы сокращения, т.е. сокращения, при которых мышца изменяет длину, относятся к динамической форме сокращения. Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим (изос - равный, метр - длина). Это статическая форма сокращения. Она возникает в двух случаях: когда внешняя нагрузка равна напряжению, развиваемому мышцей при сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют условия для растяжения мышцы под влиянием этой внешней нагрузки. При динамических формах сокращения производится внешняя работа: при концентрическом сокращении - положительная, при эксцентрическом - отрицательная. Величина работы в обоих случаях определяется как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении "расстояние" равно нулю, и, согласно физическому закону, в этом случае мышца не производит никакой работы. Однако с физиологической точки зрения изометрическое сокращение требует расхода энергии и может быть очень утомительным. В этом случае работа может быть определена как произведение величины напряжения мышцы на время ее сокращения (т.е. эквивалентно импульсу силы в физике). Во время изометрического сокращения в тепло превращается вся выделяемая мышцей энергия, а динамическом сокращении не менее 50% ее энергии. В табл. приводятся основные характеристики различных форм и типов мышечных сокращений.
В реальных условиях деятельности мышц практически не встречается чисто изометрическое или чисто изотоническое сокращение. В частности, при выполнении движений внешняя нагрузка на сокращающиеся мышцы изменяется хотя бы уже потому, что изменяются механические условия их действия (плечи действия сил и угол их приложения). Следовательно, такое сокращение мышц уже не является чисто изотоническим, т.е. с постоянным напряжением. Чисто изометрическое сокращение мышц также вряд ли возможно. В реальных условиях нельзя абсолютно жестко зафиксировать сустав, исключив изменение длины мышц. Даже в экспериментальных условиях, когда сухожильные концы мышцы строго фиксированы, напряжение собственно сократительных элементов передается упругим пассивным элементам мышцы, вызывая некоторое их растяжение. В результате происходит укорочение собственно сократительной части мышцы. Смешанную форму сокращения, при которой изменяется и Длина, и напряжение мышцы, называют ауксотонической, или анизотонической. Эта форма сокращения преобладает в натуральных условиях работы мышц в теле человека. С помощью специальных устройств можно регулировать внешнюю Нагрузку (сопротивление движению в суставе) таким образом, что с Ростом напряжения мышцы (суставного момента) сопротивление Движению в такой же мере увеличивается, а при уменьшении Мышечного напряжения - сопротивление настолько же снижается. В этом случае при постоянной (максимальной) активации, мыщц движение осуществляется с постоянной скоростью. Такой тип динамического сокращения мышц называется изокинетическим. По-видимому, при выполнении пловцом гребка рукой в воде мышцы работают в режиме, близком к изокинетическому, так как с увеличением скорости движения руки пропорционально возрастает сопротивление воды. В результате скорость перемещения руки в воде (скорость укорочения мышц) близка к постоянной.
№ 24 Работа, сила, мощность мышцы, утомление. Различают следующие режимы мышечного сокращения: Максимальная сила мышц - это величина максимального напряжения, которое может развить мышца. Она зависит от строения мышцы, ее функционального состояния, исходной длины, пола. возраста, степени тренированности человека. В зависимости от строения, выделяют мышцы с параллельными волокнами (например, портняжная'. веретенообразные (двуглавая мышца плеча), перистые (икроножная). У этих типов мышц различная площадь поперечного физиологического сечения. Это сумма площадей поперечного сечения всех мышечных волокон образующих мышцу. Наибольшая площадь поперечного физиологического сечения а, следовательно, сила, у перистых мыши. Наименьшая у мышце параллельным расположением волокон (рис.). Утомление мышц Утомление - это временное снижение работоспособности мыши в результате работы. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается (рис). Чем выше частота, сила раздражения, величина нагрузки тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, .но/снижается амплитуда (рис.) Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях, полного расслабления не наступает, развивается контрактура. Это состояние непроизвольного длительного сокращения мышцы. Работа утомление мышц исследуются с помощью эргографии. В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления.
Поскольку основной задачей скелетной мускулатуры является совершение мышечной работы, в экспериментальной и клинической физиологии оценивают величину работы, которую совершает мышца, и мощность, развиваемую ею при работе. Согласно законам физики, работа есть энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние: А = FS. Если сокращение мышцы совершается без нагрузки (в изотоническом режиме), то механическая работа равна нулю. Если при максимальной нагрузке не происходит укорочения мышцы (изометрический режим), то работа также равна нулю. В этом случае химическая энергия полностью переходит в тепловую. Согласно закону средних нагрузок, мышца может совершать максимальную работу при нагрузках средней величины. При сокращении скелетной мускулатуры в естественных условиях преимущественно в режиме изометрического сокращения, например при фиксированной позе, говорят о статической работе, при совершении движений — о динамической. Сила сокращения и работа, совершаемая мышцей в единицу времени (мощность), не остаются постоянными при статической и динамической работе. В результате продолжительной деятельности работоспособность скелетной мускулатуры понижается. Это явление называется утомлением. При этом снижается сила сокращений, увеличиваются латентный период сокращения и период расслабления. Статический режим работы более утомителен, чем динамический. Утомление изолированной скелетной мышцы обусловлено прежде всего тем, что в процессе совершения работы в мышечных волокнах накапливаются продукты процессов окисления — молочная и пировиноградная кислоты, которые снижают возможность генерирования ПД. Кроме того, нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для энергообеспечения мышечного сокращения. В естественных условиях мышечное утомление при статической работе в основном определяется неадекватным регионарным кровотоком. Если сила сокращения в изометрическом режиме составляет более 15% от максимально возможной, то возникает кислородное «голодание» и мышечное утомление прогрессивно нарастает. В реальных условиях необходимо учитывать состояние ЦНС — снижение силы сокращений сопровождается уменьшением частоты импульсации нейронов, обусловленное как их прямым угнетением, так и механизмами центрального торможения. Еще в 1903 г. И. М. Сеченов показал, что восстановление работоспособности утомленных мышц одной руки значительно ускоряется при совершении работы другой рукой в период отдыха первой. В отличие от простого отдыха такой отдых называют активным. Работоспособность скелетной мускулатуры и скорость развития утомления зависят от уровня умственной деятельности: высокий уровень умственного напряжения уменьшает мышечную выносливость.
Механическая работа, выполняемая мышцей, равна произведению развиваемой ею силы и расстояния, на протяжении которого она действует. Мощность мышечного сокращения отличается от силы мышцы, поскольку мощность является мерой общего количества работы, выполняемой в единицу времени. Следовательно, мощность определяется не только силой мышечного сокращения, но также расстоянием сокращения и числом сокращений в минуту. Мышечная мощность обычно измеряется в килограммометрах (кгм) в минуту. Например, о мышце, которая может поднимать вес, равный 1 кг, на высоту 1 м или сдвигать некий объект в сторону с силой 1 кг на расстояние 1 м за 1 мин, говорят, что ее мощность равна 1 кгм/мин. Максимальная мощность, развиваемая всеми мышцами очень тренированного спортсмена при условии их совместной работы, примерно следующая. Таким образом, ясно, что человек может развивать чрезвычайную мощность в течение коротких периодов времени, например во время забега на 100 м, который полностью завершается в течение 10 сек, тогда как для долговременных нагрузок, требующих выносливости, выходная мощность мышц составляет лишь 1/4 от первоначальной мощности. Это не означает, что спортивная производительность человека в 4 раза больше в период первичного всплеска по сравнению со следующими 30 мин, поскольку эффективность при переводе мощности мышц на производительность спортсмена часто гораздо меньше во время быстрой активности, чем во время длительной активности, но менее быстрой. Так, скорость бега на 100 м лишь в 1,75 раз выше скорости 30-минутного бега, несмотря на четырехкратную разницу допустимой мощности мышц при кратковременном беге по сравнению с длительным.
№ 25 Нейро-моторная (двигательная) единица, ее виды и функции. НЕЙРО-МОТОРНЫЕ ЕДИНИЦЫ. Структурно-функциональной единицей скелетных мышц является многоядерное мышечное волокно. Объединяясь в пучки, эти волокна образуют мышцу. Любое мышечное волокно иннервируется только от одного мотонейрона. В то же время один мотонейрон иннервирует группу мышечных волокон, вместе с которыми он образует двигательную единицу. Она является наименьшей функциональной единицей моторной системы. В разных мышцах двигательные единицы сильно отличаются по количеству мышечных волокон. Так, например, в глазо-двигательных мышцах на один нейрон приходится около десятка мышечных волокон, в двигательных единицах мелких мышц руки их около сотни, а в крупных мышцах конечностей и туловища - более тысячи на один мотонейрон. Малые единицы характерны для мышц, с помощью которых совершаются точные, хорошо координированые движения, тогда как большие преобладают в массивных мышцах, таких например, как мышцы спины. В зависимости от скорости развития максимальной силы сокращения, а также от быстроты утомления различают два типа двигательных единиц: 1) быстрый, утомляемый тип; 2)медленный, мало утомляемый. В большинстве мышц представлены два типа двигательных единиц, но в разном соотношении, которое варьирует у людей индивидуально. В спортивной физиологии людей с относительно большим процентом быстрых волокон относят к спринтерам, а обладателей большего, чем в среднем, процента медленных волокон - к стайерам. У всех людей при старении процент быстрых волокон уменьшается. Волокна быстрых двигательных единиц относительно меньше снабжаются кровью и поэтому имеют бледно-красный цвет. Они предназначены для мощных быстрых сокращений в течение короткого времени, поскольку быстро утомляются, не поспевая из-за относительно малого кровоснабжения окислять глюкозу до конечных продуктов. Волокна медленного типа обеспечены обильной сетью кровеносных капилляров, что придаёт им тёмно-красный цвет. И, хотя они сокращаются медленнее, зато способны очень долго работать без утомления. Развиваемая ими сила составляет от 1 до 10% от силы быстрых волокон. Тела мотонейронов двигательных единиц медленного типа имеют наименьшие размеры и самый низкий порог возбуждения, поэтому только они могут активироваться слабыми входными сигналами. Мотонейроны быстрых двигательных единиц, напротив, являются самыми крупными и, чтобы возбудить их, нужны сильные стимулы. Поэтому медленные двигательные единицы используются при любых движениях, а быстрые - только в тех случаях, когда понадобится большая сила сокращающихся мышц. Так, например, при стоянии или спокойной ходьбе сокращение мышц обеспечивает менее чем половина всех двигательных единиц - здесь сила мышечного сокращения составляет около 20% от максимальной. Для того чтобы выполнить прыжок, может потребоваться максимальная сила: в таком случае понадобится активация мотонейронов быстрых двигательных единиц. № 26 Строение и принцип функционирования нервно-мышечного синапса. Синапс– это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннервирующую клетку. Структура синапса: 1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке); 2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс); 3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови). В синаптическом окончании аксона находятся пузырьки (везикулы), а в них ацетилхолин.
Потенциал действия вызывает открытие кальциевых каналов и вход Са. Са вызывает движение везикул к с ацетилхолином к внутренней поверхности пресинаптической мембраны. Дальше происходит секреция медиаторов. Мембранные везикулы и пресинаптическая мембрана имеют сходное строение, и мембранные везикулы начинают встраиваться в пресинаптическую мембрану, в результате чего площадь пресинаптической мембраны увеличивается, и ацетилхолин поступает в щель. Далее он соединяется с рецепторами постсинаптической мембраны, являясь для них адекватным раздражителем, вызывает локальный ответ. В результате суммации локальных ответов, возникает потенциал действия, который может распространяться. Затем ацетилхолин разрушается ферментом ацетилхолин-эстеразой. Элементы разрушения ацетилхолина идут в тело нейрона, где ресинтезируются, которое возвращается обратно в синаптическое окончание, где упаковывается в везикулу.
№ 27 Классификация нервных волокон. Классификация нервных волокон по их морфологическим особенностям:
Примечания: Группа А делится на альфа, бета, гамма, дельта волокна. Альфа – двигательные и проприоцептивные. Бета – от сознательных рецепторов. Гамма – от температурных рецепторов. Дельта – от болевых рецепторов. В1-В2 – чувствительные волокна внутри органов, преганглионарные волокна.
Функциональная классификация периферических нервных волокон 1. Чувствительные (афферентные, или сенсорные) нервные волокна 2. Двигательные (эфферентные, или моторные) нервные волокна. Классификация периферических нервных волокон (Эрлангер Д., Гассер Г., 1937) ) Критерии классификации 1.диаметр волокна 2. возбудимость волокна 3. временные характеристики ПД волокна 4. скорость проведения возбуждения Классификация волокон Эрлангера и . Гассера • Волокна группы А – альфа, бета, гамма ( V=70-120, 40-70 и 15-40 м/с), •Волокна группы В (V=73-14 м/с) •Волокна группы С (0,5-2,0. м/с) Волокна группы А альфа (диаметр -13-22 мкм, скорость – 60- 120 м/с, длительность ПД- 0,4-0,5 мс) 1). эфферентные волокна, проводящие возбуждение к скелетным мышцам от альфа-мотонейронов 2) афферентные волокна, проводящие возбуждение от мышечных рецепторов в ЦНС Волокна группы А бета (диаметр – 8-13- мкм, скорость – 40- 70 м/с, длительность ПД- 0,4 -0,6 мс) 1. Афферентные волокна, проводящие возбуждение от рецепторов прикосновения и сухожильных рецепторов в ЦНС Волокна группы А гамма (диаметр – 4-8 мкм, скорость – 15- 40 м/с, длительность ПД- 0,5 -0,7 мс) 1) эфферентные волокна к мышечным веретенам от гамма-мотонейрнов 2). афферентные волокна, проводящие возбуждение от рецепторов прикосновения и давления в ЦНС Волокна группы В (диаметр – 1-3 мкм, скорость -3-14 м/с, длительность ПД- 1,2 мс) Это – преганглионарные волокна вегетативной нервной системы Волокна группы С (диаметр – 0,5—1,0 мкм, скорость -0,5-2,0 м/с, длительность ПД- 2,0 мс) 1.постганглионарные волокна ВНС 2.афферентные волокна, проводящие возбуждение от рецепторов боли, давления и тепла в ЦНС
№ 28 Особенности строения миелиновых и безмиелиновых нервных волокон. Типы нервных волокон: - безмякотный – безмиелиновый; - мякотный – миелиновый. Миелин – вещество белково-липидной природы, обладающее свойствами диэлектрика, вырабатывается в клетках Шванна, которые спиралеобразно накручены на осевой цилиндр. Перехваты Ранвье – участки волокна е покрытые миелиновой оболочкой. Скачкообразный или сальтоторный способ передачи возбуждения на миелиновом волокне – возбуждение перескакивая через участок покрытый миелином от одного перехвата Ранвье к другому. Чем выше и толще волокно, тем больше скорость распространения возбуждения, и тем меньше тратиться энергии.
Физиологические свойства и функциональная значимость нервных волокон Нервные волокна имеют самую высокую возбудимость, самую высокую скорость проведения возбуждения, самый короткий рефрактерный период, высокую лабильность. Это обеспечивается высоким уровнем обменных процессов и низкой величиной мембранного потенциала. Функция: проведение нервных импульсов от рецепторов к центральной нервной системе и обратно. Особенности строения и виды нервных волокон Нервное волокно - аксон - покрыт клеточной мембраной. |