| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Структурно-химическая характеристика мембран клеток
Биологическая мембрана Общей чертой всех мембран клетки является то, что они представляют собой тонкие (6—10 нм) пласты липопротеидной природы (т.е. липиды в комплексе с белками). Основными химическими компонентами клеточных мембран являются липиды (~40%), белки (~60%) и углеводы (5—10%). К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и растворимостью в органических растворителях и жирах (липофильность). Состав липидов очень разнообразен. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды (глицерофосфатиды), сфингомиелины и из стероидных липидов — холестерин. От холестерина зависит текучесть и стабильность мембран. Особенностью липидов мембран является разделение их молекул на две функционально различные части: гидрофобные неполярные, не несущие зарядов «хвосты», состоящие из жирных кислот, и гидрофильные, заряженные полярные «головки». Это определяет способность липидов самопроизвольно образовывать двухслойные (т.е. билипидные) мембранные структуры толщиной 5—7 нм. Различные клеточные мембраны могут значительно отличаться друг от друга по липидному составу и набору белковых молекул. Многие мембранные белки состоят из двух частей, из участков, богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными аминокислотами: глицином, аланином, валином, лейцином. Такие белки в липидных слоях мембран располагаются так, что их неполярные участки как бы погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная) же часть этих белков взаимодействует с головками липидов и обращена в сторону водной фазы. Среди белков клеточной мембраны выделяют т.н. интегральные белки, пронизывающие ее насквозь, и примембранные, или поверхностные, не встроенные в билипидный слой. По биологической роли белки мембран можно разделить на белки-ферменты, белки-переносчики, рецепторные и структурные белки. Углеводы мембран связаны с молекулами липидов или белков. Такие вещества называются соответственно гликолипидами и гликопротеинами. Количество их в мембранах обычно невелико. Как бы ни было велико различие между мембранами по количеству и составу их липидов, белков и углеводов, мембраны обладают рядом общих свойств, определяемых их основной структурой. Все мембраны являются барьерными структурами, резко ограничивающими свободную диффузию веществ между цитоплазмой и средой, с одной стороны, и между гиалоплазмой и содержимым мембранных органелл — с другой. Особенность же специфических функциональных нагрузок каждой мембраны определяется свойствами и особенностями белковых компонентов, большая часть из которых представляет собой ферменты или ферментные системы. Большую роль в функционировании мембран играют гликолипиды и гликопротеиды.
Рис. 1. Строение клеточной мембраны (схема). 1 — липиды; 2 — гидрофобная зона бислоя липидных молекул; 3 — интегральные белки мембраны; 4 — полисахариды гликокаликса.
Плазмолемма (plasmalemma), или внешняя клеточная мембрана, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку. Химический состав плазмолеммы. Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран. Снаружи от плазмолеммы располагается надмембранный слой — гликокаликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы (рис. 1). При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) видно, что они образуют как бы чехол поверх плазматической мамбраны. В гликокаликсе могут располагаться белки, не связанные непосредственно с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др. Функции плазмолеммы Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются барьерная функция (разграничения цитоплазмы с внешней средой), функции рецепции и транспорта различных веществ как внутрь клетки, так и из нее. Рецепторные функции связаны с локализацией на плазмолемме специальных структур, участвующих в специфическом «узнавании» химических и физических факторов. Клеточная поверхность обладает большим набором компонентов — рецепторов, определяющих возможность специфических реакций с различными агентами. Рецепторами на поверхности клетки могут служить гликопротеиды и гликолипиды мембран (рис. 1). Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны. Существуют рецепторы к биологически активным веществам — гормонам, медиаторам, к специфическим антигенам разных клеток или к определенным белкам. С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток, развитие иммунитета, рецепторов, реагирующих на физические факторы. Так, в плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что в свою очередь приводит к генерации электрического импульса.
Транспорт веществ Выполняя транспортную функцию, плазмолемма обеспечивает диффузию (пассивный перенос) ряда веществ, например воды, ионов, некоторых низкомолекулярных соединений. Другие вещества проникают через мембрану путем активного переноса против градиента концентрации с затратой энергии за счет расщепления АТФ. Так транспортируются многие органические молекулы (сахара, аминокислоты и др.). Эти процессы могут быть сопряжены с транспортом ионов, в них принимают участие специальные белки-переносчики.
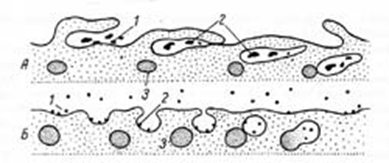
Рис. 2. Эндоцитоз. Разные типы (А, Б) образования пиноцитозных пузырьков. 1 — сорбция частиц на поверхности плазматической мембраны; 2 — погружение частиц в цитоплазму; 3 — первичные лизосомы.
Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процессов эндоцитоза (рис. 2, А, Б). Эндоцитоз формально разделяют на фагоцитоз (захват и поглощение клеткой крупных частиц, например бактерий или даже фрагментов других клеток), и пиноцитоз (захват макромолекулярных соединений). Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Эти впячивания могут иметь вид еще незамкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней. В дальнейшем эндоцитозные пузырьки могут сливаться друг с другом, расти и в их внутренней полости, кроме поглощенных веществ, начинают обнаруживаться гидролитические ферменты (гидролазы), поступающие сюда из лизосом. Эти ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиа-лоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению. Плазмолемма принимает участие в выведении веществ из клетки (т.н. экзоцитоз). В этом случае внутриклеточные продукты (белки, мукополисахариды, жировые капли и др.), заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются и содержимое вакуоли поступает в окружающую среду. Процесс эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы — таких, как микротрубочки и сократимые микрофиламенты. Последние, соединяясь с определенными участками плазмолеммы, могут, изменяя свою длину, втягивать мембрану внутрь клетки, что приводит к отделению от плазмолеммы эндоцитозных вакуолей. Часто, непосредственно примыкая к ней, микрофиламенты образуют сплошной, так называемый кортикальный слой. Плазмолемма многих клеток животных может образовывать выросты различной структуры. У ряда клеток такие выросты включают в свой состав специальные компоненты цитоплазмы (микротрубочки, микрофибриллы), что приводит к развитию специальных структур — микроворсинок, ресничек, жгутиков. Наиболее часто встречаются на поверхности многих животных клеток микроворсинки. Эти выросты цитоплазмы, ограниченные плазмолеммой, имеющие форму цилиндра с закругленной вершиной. Микроворсинки характерны для клеток эпителиев, но обнаруживаются и у клеток других тканей. Диаметр микроворсинок около 100 нм. Число и длина их различны у разных типов клеток. Возрастание числа микроворсинок приводит к резкому увеличению площади клеточной поверхности. Это особенно важно для клеток, участвующих во всасывании. Так, в кишечном эпителии на 1 мм2 поверхности насчитывается до 2•10^8 микроворсинок.
2. Сперматогенез Образование мужских половых клеток (сперматогенез) протекает в извитых семенных канальцах и включает 4 последовательные стадии или фазы: размножение, рост, созревание и формирование. Начальной фазой сперматогенеза является размножение сперматогоний, занимающих наиболее периферическое (базальное) положение в сперматогенном эпителии. Согласно современным представлениям, среди сперматогоний можно выделить два типа клеток: 1) стволовые сперматогонии типа А, которые подразделяются на две субпопуляции: долгоживущие, резервные стволовые клетки и быстро обновляющиеся полустволовые клетки, которые делятся один раз в течение цикла сперматогенного эпителия, 2) дифференцирующиеся сперматогоний типа А и типа В. Стволовые клетки расположены в базальной части канальца изолированно от других сперматогоний. Морфологически в популяции стволовых А-сперматогоний различают светлые и темные клетки. Для обоих клеток характерно преобладание в ядрах деконденсированного хроматина и расположение ядрышек около ядерной оболочки. Однако в темных клетках типа А степень конденсации хроматина большая, чем в светлых клетках. Темные клетки относят к «резервным» медленно обновляющимся стволовым клеткам, а светлые — к полустволовым быстро обновляющимся клеткам. Для стволовых клеток характерно наличие овальных ядер с диффузно распределенным хроматином, одного или двух ядрышек, большое содержание в цитоплазме рибосом и полисом, малое количество других органелл. Часть стволовых клеток типа А при делении не завершают цитокинез и остаются соединенными цитоплазматическими мостиками, т.е. образуют синцитий. Появление таких спаренных сперматогоний свидетельствует о начале процессов дифференцировки мужских половых клеток. Дальнейшее деление таких клеток приводит к образованию цепочек или групп сперматогоний, соединенных цитоплазматическими мостиками. Клетки типа В имеют более крупные ядра, хроматин в них не дисперсный, а собран в глыбки. В следующий период (период роста) сперматогонии перестают делиться митозом, увеличиваются в объеме и вступают в первое деление мейоза. Это начало их дифференцировки в сперматоциты 1-го порядка, и начало уже третьего периода – периода созревания. Синцитиальные группы сперматогоний начинают перемещаються в адлюминальную зону сперматогенного эпителия. В первом делении мейоза в клетках происходит конъюгация гомологичных хромосом и кроссинговер. В каждой из двух дочерних клеток — сперматоцитов 2-го порядка содержится гаплоидное число хромосом (23 у человека). Второе деление созревания начинается сразу вслед за первым, и происходит как обычный митоз без редупликации хромосом. В анафазе второго деления созревания диады сперматоцитов второго порядка разъединяются на монады, или одиночные хроматиды, расходящиеся к полюсам. В результате сперматиды получают столько же монад, сколько было диад в ядрах сперматоцитов второго порядка, т.е. гаплоидное число. Сперматоциты 2-го порядка имеют меньшие размеры, чем сперматоциты 1-го порядка, и располагаются в средних и более поверхностных слоях эпителиосперматогенного слоя. Таким образом, каждая исходная сперматогония дает начало 4 сперматидам с гаплоидным набором хромосом. Сперматиды больше не делятся, но путем сложной перестройки превращаются в зрелые сперматозоиды. Эта трансформация составляет четвертую фазу сперматогенеза — период формирования, или спермиогенеза. Сперматиды представляют собой небольшие округлые клетки со сравнительно крупными ядрами. Скапливаясь около верхушек поддерживающих клеток, сперматиды частично погружаются в их цитоплазму, что создает условия для формирования сперматозоидов из сперматид. Ядро сперматиды постепенно уплотняется и уплощается. В сперматидах около ядра располагаются аппарат Гольджи, центросома и скапливаются мелкие митохондрии. Процесс формирования сперматозоида начинается с образования в зоне аппарата Гольджи уплотненной гранулы — акробласта, прилежащего к поверхности ядра. В дальнейшем акробласт, увеличиваясь в размерах, в виде шапочки охватывает ядро, а в середине акробласта дифференцируется уплотненное тельце. Такую структуру называют акросомой. Она лежит в том конце трансформирующей сперматиды, который обращен к поддерживающей клетке. Центросома, состоящая из двух центриолей, перемещается в противоположный конец сперматиды. Проксимальная центриоль прилегает к поверхности ядра, а дистальная разделяется на две части. От передней части дистальной центриоли начинает формироваться жгутик (flagellum), который затем становится осевой нитью развивающегося сперматозоида. Задняя же половина дистальной центриоли принимает вид колечка. Смещаясь вдоль жгутика, это колечко определяет заднюю границу средней или связывающей части сперматозоида. Цитоплазма по мере роста хвоста сползает с ядра и сосредоточивается в связующей части. Митохондрии располагаются спиралеобразно между проксимальной центриолью и колечком. Цитоплазма сперматиды во время ее превращения в спермий сильно редуцируется. В области головки она сохраняется только в виде тонкого слоя, покрывающего акросому; небольшое количество цитоплазмы остается в области связующего отдела и, наконец, она очень тонким слоем покрывает жгутик. Часть цитоплазмы сбрасывается и распадается в просвете семенного канальца либо поглощается поддерживающими клетками. Кроме того, эти клетки вырабатывают жидкость, накапливающуюся в просвете извитого семенного канальца. В эту жидкость попадают, высвобождаясь из верхушек поддерживающих клеток, сформировавшиеся сперматозоиды и вместе с ней уходят в дистальные части канальца. Процесс сперматогенеза в целом длится у человека около 75 сут, но протекает на протяжении извитого семенного канальца волнообразно. Поэтому на каждом отрезке канальца имеется определенный набор клеток сперматогенного эпителия. Эпителиосперматогенный слой чрезвычайно чувствителен к повреждающим действиям. При различных интоксикациях, авитаминозах, недостаточности питания и других условиях (особенно при воздействии ионизирующим излучением) сперматогенез ослабляется или даже прекращается, а сперматогенный эпителий атрофируется. Аналогичные деструктивные процессы развиваются при крипторхизме (когда семенники не опускаются в мошонку, оставаясь в брюшной полости), длительном пребывании организма в среде с высокой температурой, лихорадочных состояний и особенно после перевязки или перерезки семявыводящих каналов. Деструктивный процесс при этом поражает в первую очередь формирующиеся сперматозоиды и сперматиды. Последние набухают, нередко сливаются в характерные округлые массы — так называемые семенные шары, плавающие в просвете канальца. Так как нижние слои сперматогенного эпителия (сперматогонии и сперматоциты 1-го порядка) при этом сохраняются более длительно, то восстановление сперматогенеза после прекращения действия повреждающего агента иногда оказывается возможным. Деструкция, описанная выше, ограничивается только сперматогенным слоем. Поддерживающие клетки в указанных обстоятельствах сохраняются и даже гипертрофируются, а гландулоциты часто увеличиваются в количестве и образуют большие скопления между запустевающими семенными канальцами. |