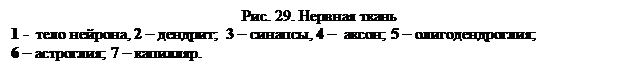| ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Игровые автоматы с быстрым выводом Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Классификация мышечных тканей
Для всей группы мышечных тканей характерна удлиненная структура их компонентов и наличие специальных органелл – миофибрилл. Сокращение мышечных тканей происходит за счет сокращения миофибрилл. Миофибриллы состоят из миофиламентов – двух типов белков: актина и миозина.
Молекула миозина состоит из головки и стержня. Головка миозина прилегает к молекуле актина но на связана с ней. В расслабленной мышце тропомиозин прикрывает химически активные участки молекул актина и тем самым лишает миозин возможности взаимодействовать с актином.
Гладкая мышечная ткань состоит из мышечных клеток – миоцитов. Миоциты имеют веретенообразную форму, длина их варьирует от 20 до 500 мкм, диаметр 5-8 мкм. Удлиненное с конденсированным хроматином ядро расположено в центре клетки, органоиды лежат на периферии. В периферической части цитоплазмы расположены миофиламенты – миозиновые и актиновые. Миозиновые миофиламенты более толстые и лежат в цитоплазме продольно. Актиновые филаменты тонкие, прикрепляются к цитолемме в области электронноплотных телец. При сокращении происходят взаимодействие молекул актина и миозина, смещение актиновых и миозиновых миофиламентов навстречу друг к другу – образование актино-миозиновых комплексов. Затем происходит фаза расслабления – распад актино-миозиновых комплексов. Ионы Ca++ поступают в клетку из каналов ЭПС. Гладкая мышечная ткань сокращается ритмично, мало утомляема и сокращается под контролем вегетативной нервной системы, образует мышечные слои внутренних органов, сосудов. Миоциты эпидермального происхождения представлены миоэпителиальными клетками. Миоэпителиоциты расположены на базальной мембране секреторных отделов молочных, потовых, сальных и слюнных желез, имеют отростчатую форму. Отростки миоэпителиоцитов охватывают секреторные клетки, и их сокращение способствует выведению секрета. Миоциты нейрального происхождения расположены в радужке глаза, образуя циркулярный (мышца, суживающая зрачок) и радиальный (мышца, расширяющая зрачок) слои. Поперечно-полосатые (исчерченные) мышечные ткани объединяют сердечную и скелетную мышечные ткани по признаку наличия в них исчерченных миофибрилл, которые обладают большей скоростью сокращения, чем гладкие миоциты. Скелетная поперечно-полосатая мышечная ткань состоит из мышечных волокон, длина которых может достигать 30 см. Развиваются волокна из миотомов, в процессе дифференцировки которых образуется два типа структур – миосимпласты и миосателлиты. Мышечные волокна – миосимпласты – покрыты сарколеммой, образованной цитоплазматической и базальной мембранами. В центре волокна лежат пучки миофибрилл. В пучках различают светлые (изотропные) I-диски, образованные актиновыми филаментами и темные (анизотропные) А-диски, состоящие из миозина. Смещение актиновых нитей относительно друг друга не происходит, поскольку через середину каждого изотропного диска проходит телофрагма и закрепляется на сарколемме. Телофрагма обеспечивает постонство положения актиновых нитей в пучке относительно друг друга. Сходная структура присутствует в А-дисках. При сокращении миофибриллы происходят те же конформационные изменения актина и миозина что при сокращении в гладкой мускулатуре, но так как в миофибриле этих волокон много и они упорядочены сокращение происходит более активно.
Клеточные органеллы: митохондрии, ядра, остатки комплекса Гольджи находятся на периферии волокна. Гладкая ЭПС в виде манжетки одевает пучки миофибрилл и соединяется с цитоплазматической мембраной сарколеммы. При передаче возбуждения с нервного окончания на мышцу происходит увеличение проницаемости мембраны ЭПС, ионы кальция выходят в саркоплазму, обуславливая сокращение волокна. Поперечно-полосатая мышечная ткань сокращается энергично, под контролем сознания, мышца утомляема. Миосателлиты– это одноядерные клетки, окружающие миосимпласт. Эти клетки являются малодифференцированными. За счет миосателлитов происходит регенерация мышечной ткани. Сердечная мышца состоит из различных типов сердечных миоцитов (кардиомиоцитов) – сократительных, проводящих и секреторных. Основной структурной единицей миокарда являются сократительные кардиомиоциты, имеющие удлиненную форму (100-150 мкм), близкую к цилиндрической. Их концы соединяются друг с другом так, что цепочки кардиомиоцитов составляют так называемые функциональные волокна толщиной 10-20 мкм, а области контакта образуют вставочные диски. Их боковые поверхности покрыты базальной мембраной, в которую снаружи вплетаются тонкие ретикулярные и коллагеновые волокна. Кардиомиоциты объединены многочисленными щелевыми клеточными контактами (анастомозами) в единую сеть. В центре кардиомиоцита находится ядро овальной формы. Сердечная мышца богата митохондриями, в цитоплазме между митохондриями часто встречаются гранулы гликогена. Эндоплазматический ретикулюм кардиомиоцитов развит не так сильно, как в скелетной мускулатуре.
Проводящие кардиомиоциты образуют проводящую систему сердца (синусно-предсердный узел, предсердно-желудочковый узел, мышечные волокна Пуркинье), от которых импульсы передаются на рабочие сократительные кардиомиоциты. Цитоплазма проводящих кардиомиоцитов окрашивается бледнее сократительных миоцитов, богата гликогеном и митохондриями, миофибриллы малочисленны и не образуют общей поперечной исчерченности. Секреторные кардиомиоциты расположены в предсердиях и содержат в цитоплазме секреторные гранулы, содержащие вещества регулирующие артериальное давление (натрийуретический фактор и др.) Сердечная мышца сокращается ритмично, она неутомляема, ее сокращения непроизвольны.
Глава 5. НЕРВНАЯ ТКАНЬ Нервная ткань является основным структурным компонентом нервной системы. Нервная ткань выполняет следующие функции: 1. осуществляет объединение (интеграцию) частей организма в единое целое; 2. обеспечивает регуляцию и корреляцию деятельности тканей и органов, 3. осуществляет взаимодействие и связь организма с окружающей средой; 4. интегративная функция ткани лежит в основе сознания, речи, мышления, памяти, эмоций, поведения. Нервная ткань воспринимает многообразную информацию, поступающую из внешней среды и из внутренних органов, перерабатывает ее и генерирует сигналы, обеспечивающие ответные реакции, адекватные действующим раздражителям. Чтобы понять гистологическое строение нервной системы, необходимо знать ее развитие в онтогенезе. Формирование нервной системы начинается с уплотнения дорсальной части эктодермы – закладки нервной пластинки, а затем идет образование нервной борозды и нервной трубки. Нервная ткань состоит из нервных клеток (нейронов) и связанных с ними клеток нейроглии. В совокупности эти структурные элементы составляют единую морфологическую и функциональную основу всех органов центральной и периферической нервной системы. Основная структурная и функциональная единица нервной ткани – нейрон – клетка способная воспринимать раздражение, приходить в состояние возбуждения, обрабатывать информацию, вырабатывать и передавать импульс. По функциональному значению нервные клетки делят: § рецепторные(афферентные) или чувствительные; § эффекторныеили эфферентные – передающие импульс на рабочий орган, и § вставочные или ассоциативные – осуществляющие связь между нейронами. Для нейрона характерно наличие отходящих от тела клетки тонких цитоплазматических отростков. По строению и функции выделяют два типа отростков: § аксон или нейрит, обычно проводящий нервный импульс от тела нервной клетки. Аксон всегда один, не ветвится, но от него могут перпендикулярно отходить коллатерали, принимающие на некотором расстоянии параллельное нейриту направление; § дендрит, он ветвится и проводит нервный импульс к телу клетки. Дендритов может быть много. По числу отростков нервные клетки делятся на униполярные с одним отростком, чаще аксоном, биполярные– с двумя отростками – аксоном и дендритом и мультиполярные – с одним аксоном и несколькими дендритами. Разновидностью биполярных нейронов являются ложноуниполярные нейроны спинального ганглия. От их тела отходит один отросток, который Т-образно делится на аксон и дендрит. Большинство нейронов человеческого организма мультиполярны, имеют пирамидную, звездчатую, грушевидную и другие формы. Размеры тела нейрона варьируют от 4 мкм (зернистые клетки) до 135 мкм (крупные пирамидные клетки Бэца).
Вторым характерным компонентом цитоплазмы нейрона являются нейрофибриллы, которые проявляются только при импрегнации солями серебра микропрепаратов нервной ткани* (* – отмечены данные по особенностям методов окраски препаратов, которые приведены в конце пособия). При электронной микроскопии было обнаружено, что нейрофибриллы – слипшиеся под действием солей серебра пучки нейротрубочек и нейрофиламентов. Нейрофиламенты (10 нм) - промежуточные филаменты. Нейротрубочки (24 нм) поддерживают форму нейрона и участвуют в транспорте нейромедиатора и других веществ. В цитоплазме тела нейрона хорошо развит комплекс Гольджи, лежащий около ядра и участвующий в синтезе нейромедиатора. Мощно развитый митохондриальный ретикулюм расположен в теле нейрона и отростках. В телах нейронов содержится два пигмента – липофусцин желто-коричневого цвета – пигмент «старения» (продукт «изнашивания» нервных клеток, неподдающийся лизосомному перевариванию) и меланин коричневого цвета, значение которого в нервной ткани неизвестно. Аксон имеет диаметр 1-20 мкм, содержит много нитевидных митохондрий, пузырьки гладкой ЭПС, нейротрубочки и нейрофиламенты. Рибосомы встречаются очень редко. Аксон покрыт олигодендроцитами. Цитоплазма в аксоне находится в движении, обеспечивая аксонный транспорт. Различают медленный ток (со скоростью 1-3 мм в сутки), с ним двигаются лизосомы, ферменты от тела клетки и быстрый ток (5-10 мм в час), с которым от тела клетки двигаются митохондрии и пузырьки нейромедиатора.
В этом процессе основную роль играют элементы цитоскелета. Различают также обратный, ретроградный (или обратный) ток, транспортирующий к телу клетки цитоплазматические компоненты со скоростью быстрого тока – 3 мм в час. Дендриты короче аксонов и обладают дихотомическим ветвлением. У нейрона имеется от 5 до 15 крупных дендритов, что составляет примерно 80-90% его поверхности, что обеспечивает возможность многочисленных связей между нейронами (10100 связей). В отличие от аксонов цитоплазма дендрита содержит гранулярную ЭПС. Передача нервного импульса осуществляется деполяризацией мембраны нейрона. В клетке, генерирующей электрические импульсы, присутствуют два типа потенциалов: потенциал действия и потенциал покоя. Плазмолемма нейрона в состоянии покоя непроницаема для ионов натрия. В результате более высокой концентрации ионов натрия у наружной стороне мембраны, она заряжается положительно, внутренняя же сторона ее имеет отрицательный заряд. Потенциал покоя плазмолеммы нейрона составляет примерно 70 мВ. На месте приложения стимула мембрана становится проницаемой для ионов натрия, ионы поступают внутрь клетки и заряд мембраны становится нейтральным или слабо положительным. Мембрана становится деполяризованной – потенциал действия. Деполяризация одного участка плазмолеммы вызывает деполяризацию соседнего участка. Нервный импульс «пробегает» по дендритам к телу клетки. В нервной ткани выделяют нейроглию, которую до недавнего времени считали вспомогательной тканью, «золушкой нервной ткани». Все клетки нейроглии делятся на макроглию и микроглию. Макроглия развивается из нервной трубки. К ней относят астроглию, эпендимоглию и олигодендроглию.
Астроглия бывает плазматической и волокнистой. Плазматические астроциты – мелкие клетки, имеющие тонкие цитоплазматические отростки, расположены в сером веществе головного и спинного мозга. Между клетками астроглии лежат нейроны. Плазматическая астроглия выполняет опорную функцию, участвует в питании нейронов и осуществлении их проводящей функции. Волокнистая астроглия входит в состав белого вещества спинного и головного мозга, выполняя опорную (для нервных волокон) функцию. Волокнистые астроциты имеют длинные слабо или неветвящиеся отростки с большим количеством фибриллярных структур. Олигодендроглия состоит из клеток малоотростчатой формы, находящихся в составе центральной и периферической нервной системы. Существует три типа олигодендроцитов: крупные светлые клетки образуются из олигодендробластов. В результате последнего деления олигодендробластов возникают светлые олигодендроциты. Они имеют крупное светлое ядро и обильную цитоплазму с множеством рибосом и других органоидов. Продолжительность жизни светлых олигодендроцитов несколько недель – постепенно они превращаются в промежуточные, а еще через несколько недель в темные олигодендроциты. В тканях взрослого организма темные олигодендроциты видны как мелкие клетки с темным крупным ядром. Они участвуют в питании нервных клеток и в проведении нервного импульса, образуя шванновскую оболочку нервных волокон. Выделяют мантийные глиоциты – олигодендроциты, окружающие тела нейронов в нервных ганглиях; нейролеммоциты, образующие шванновские оболочки нервных волокон и локализованные в нервных окончаниях терминальные глиоциты. Эпендимоглия выстилает спинномозговой канал и желудочки головного мозга. Эпендимоциты лежат подобно однослойному эпителию, выполняя разграничительную функцию. От базальной части клетки отходят отростки, проходящие в серое вещество, выполняя опорную функцию. Апикальный конец несет реснички, биение которых создает ток спинномозговой жидкости. Эпендимоциты участвуют в синтезе некоторых компонентов спинномозговой жидкости. Клетки микроглии – мелкие отростчатые, подвижные макрофаги, развивающиеся из моноцитов. Выполняют защитную (фагоцитарную) функцию. Тела нервных клеток образуют серое вещество головного и спинного мозга и нервные ганглии. Связь ЦНС с органами осуществляется при помощи проводящих элементов – нервов, основу которых составляют нервные волокна. Главную часть нервного волокна образует осевой цилиндр, представляющий собой отросток нейрона. Осевой цилиндр одет шванновской оболочкой. По строению волокна подразделяются на мякотные(миелиновые) и безмякотные (безмиелиновые). Безмякотные нервные волокна сравнительно просто построенные нервные проводники, идущие к внутренним органам. Состоит безмякотное волокно из 7-12 осевых цилиндров, идущих внутри тяжа цепочки шванновских клеток. Эти клетки многоядерные с хорошо развитой ЭПС, комплексом Гольджи и митохондриями. Безмякотное волокно образуется путем вдавления осевых цилиндров в шванновские клетки, при этом цитоплазма и плазмолемма последних охватывает осевой цилиндр и соединяется над ним. В месте их соединения образуется двойная мембрана – мезаксон. Отросток и шванновская оболочка разделены. Мякотные волокна характеризуются очень быстрым и точным проведением нервных импульсов, что обеспечивается хорошей изоляцией. Мякотные волокна идут к чувствительным и двигательным нервным окончаниям. Мякотное волокно образуется путем «накручивания» цепочки шванновских клеток на отросток нервной клетки, поэтому мякотное волокно содержит один осевой цилиндр. Концентрически закрученный вокруг осевого цилиндра мезаксон формирует миелиновый слой, образованный из чередующихся слоев белка и липидов плазматической мембраны клеток Шванна. Наружная часть шванновской оболочки представлена телом нейролеммоцита. В местах соединения шванновских клеток миелиновый слой отсутствует – эти участки называются перехваты Ранвье. В миелиновых волокнах происходит сальтаторное(от лат. прыгать)проведение нервного импульса, так как возникновение деполяризации мембраны возможно только на перехватах Ранвье. А между перехватами деполяризация быстро распространяется по осевому цилиндру (как будто перепрыгивает). Нервные окончания – это концевые аппараты отростков нейронов. Различают рецепторные или чувствительные и эффекторные нервные окончания. Рецепторные нервные окончания образованы концевыми разветвлениями дендритов чувствительных клеток. По функции различают экстерорецепторы, воспринимающие раздражение из окружающей среды и интерорецепторы, по которым сигналы идут от внутренних органов. По строению различают свободные и несвободные нервные окончания. Свободно-лежащие нервные окончания состоят из терминального разветвления дендрита, утратившего глиальную оболочку, расположены в коже и являющиеся болевыми рецепторами. Несвободные нервные окончания подразделяются на неинкапсулированные и инкапсулированные. Несвободные неинкапсулированные нервные окончания образованы терминальным разветвлением дендрита, покрытым скоплением глиоцитов – внутренней колбой.
Несвободные инкапсулированные нервные окончания кроме терминалей нервного волокна и глиоцитов содержат соединительно-тканную капсулу – наружную колбу. К инкапсулированным окончаниям относят тельце Фатера-Пачини, воспринимающее давление, колбу Краузе обеспечивающую температурное восприятие и тактильное тельце Мейсснера. К инкапсулированным чувствительным нервным окончаниям относятся так же нервно-мышечные веретена, фиксирующие изменение длины мышцы и нервно-сухожильные веретена, определяющие напряжение сухожилия. Эфекторные нервные окончания бывают двух типов: двигательные и секреторные. Двигательные нервные окончания образованы концевыми разветвле-ниями аксонов двигательных нейронов. Двигательные нервные окончания на поперечнополосатых мышечных волокнах называются моторными бляшками. Миелиновое нервное волокно подходит к мышечному, терминальная веточка аксона вдавливается в мышечное волокно, прогибая его плазмолемму. В этом участке аксон утрачивает глиоциты, мембрана которых переходит в базальную мембрану сарколеммы. Оболочка аксона не контактирует с плазмолеммой мышечного волокна, между ними имеется синаптическая щель, терминаль аксона образует пресинаптический полюс, а участок мышечного волокна – постсинаптический. Терминаль содержит синаптические пузырьки, заполненные нейромедиатором. Передача нервного импульса на мышцу подобна механизму химической синаптической передачи. В передаче информации между нервными клетками участвует специальный тип клеточных контактов – синапсы.По типу взаимодействия синапсы подразделяются на химические и электрические. В химических синапсах взаимодействие осуществляется посредством специального вещества – медиатора, тогда как в электрических синапсах для передачи импульса необходимо специфическое перераспределение токов. Большинство синапсов являются химическими и являются утолщением терминали аксона одного нейрона на дендрите, теле или аксоне другого нейрона. По локализации различают асксо-дендритные, аксо-аксонные и аксо-соматические синапсы. Терминальное утолщение аксона образует пресинаптическую часть синапса, а ее оболочка – пресинаптическую мембрану. Пресинаптическая мембрана имеет специфическое строение – образована чередующимися в шахматном порядке более толстыми и более тонкими участками плазмолеммы. В пресинаптической части находятся пузырьки с нейромедиатором (ацетилхолин, норадреналин и др.). Часть синаптических пузырьков лежит в области тонких участков мембраны. Контактирующая часть второго нейрона называется постсинаптической, а ее оболочка – постсинаптической мембраной. Между пресинаптической и постсинаптической мембранами находится синаптическая щель. Химическая синаптическая передача характеризуется высвобождением медиатора в синаптическую щель, его диффузией и последующим взаимодействием со специфическими рецепторами. Образование комплекса медиатор-рецептор вызывает открытие ионных каналов. Проходящие через них ионные токи изменяют мембранный потенциал второго нейрона. Список использованной литературы: 1. Атлас сканирующей электронной микроскопии клеток, тканей и органов под ред. О.В.Волковой, В.А.Шахламова, А.А.Миронова //М,, Медицина 1987 2. А.Афанасьев, Н.А. Юрина Гистология // М. Медицина 1989 3. Г. Л. Билич, В. А. Крыжановский Биология Полный курс. Анатомия // Издательский дом “ОНИКС 21 век” 2002 4. В.Л.Быков Частная гистология человека// Санкт-Петербург, Сотис, 1997 5. О.В.Волкова и Ю.К.Елецкий Основы гистологии с гистологической техникой // М., Медицина, 1981 6. Волкова О.В., Пекарский М.И. Эмбригионез и возрастная физиология внутренних органов человека. // М., Медицина, 1976 7. Г.Елисеев, Ю.А.Афанасьев, Е.Ф.Котовский Атлас микроскопического и ультрамикроскопического строения клеток, тканей и органов.// М., Медицина ,1970 8. А.А. Заварзин Основы частной цитологии и сравнительной гистологии многоклеточных животных // Л. Наука 1976 9. Б. Карлсон Основы эмбриологии по Пэттену // М. Мир 1983 10. Лабораторные занятия по курсу гистологии, цитологии и эмбриологии под ред. Ю.И. Афанасьева // М.. Высшая школа, 1990. 398 с. 11. Международная гистологическая номенклатура. Под ред. В.В. Семченко, Р.П. Самусева, М.В. Моисеева, З.Л. Колосовой // Омск Омская медицинская академия, 1999. 12. Практикум по гистологии, цитологии и эмбриологии. под ред. Н.А.Юриной, А.И.Радостиной // М.. Издательство Университета дружбы народов, 1989. 13. А. Ройт Основы эмбриологии // М. Мир 1991 14. Р.П. Самусев Н.И. Гончаров Эпонимы в морфологии // М. Медицина 1989. 15. И. Станек Эмбриология человека // Братислава Веда 1977. 16. Хэм А., Кормак Д. Гистология // М., Мир, 1983 в 5 томах. 17. Ю.С.Ченцов Общая цитология. //Издательство Московского университета, 1984 18. Н.А. Юрина, А.И.Радостина Гистология // М., Медицина, 1995 Список рекомендованной литературы · А.Афанасьев, Н.А. Юрина Гистология // М. Медицина 1989 · Г. Л. Билич, В. А. Крыжановский Биология Полный курс. Анатомия // Издательский дом “ОНИКС 21 век” 2002 · В.Л.Быков Частная гистология человека// Санкт-Петербург, Сотис, 1997 · О.В.Волкова и Ю.К.Елецкий Основы гистологии с гистологической техникой // М., Медицина, 1981 · Волкова О.В., Пекарский М.И. Эмбригионез и возрастная физиология внутренних органов человека. // М., Медицина, 1976 · Б. Карлсон Основы эмбриологии по Пэттену // М. Мир 1983 · Грин, Тейлор Стаут Биология // М. Мир 1991
Бессолицына Екатерина Андреевна, к.б.н., Рисунки - Казьмин Андрей Алексеевич, просто лось.
|




 Для понимания природы мышечного сокращения в 1954 году Альбертсом была предложена модель скользящих нитей: две цепочки глобулярных молекул актина обвивают друг друга и образуют двойную спираль – остов филамента. В продольных бороздах, тянущихся вдоль актинового филамента, лежат молекулы тропомиозина, которые придают актиновому филаменту жесткость.
Для понимания природы мышечного сокращения в 1954 году Альбертсом была предложена модель скользящих нитей: две цепочки глобулярных молекул актина обвивают друг друга и образуют двойную спираль – остов филамента. В продольных бороздах, тянущихся вдоль актинового филамента, лежат молекулы тропомиозина, которые придают актиновому филаменту жесткость. К молекуле тропомиозина присоединены молекулы тропонина. К актиновому филаменту прилегает миозин.
К молекуле тропомиозина присоединены молекулы тропонина. К актиновому филаменту прилегает миозин.




 Ядро нейрона крупное, сферической формы с деконденсированным хроматином. Крупные одно или несколько ядрышек на светлом фоне ядра хорошо видны (какой–то исследователь сравнил такое необычное ядро с глазом совы). Некоторые нейроны являются типичными тетраплоидными. Для нейрона характерно наличие специфических структур: нейрофибрилл и хроматофильной субстан-ции.
Ядро нейрона крупное, сферической формы с деконденсированным хроматином. Крупные одно или несколько ядрышек на светлом фоне ядра хорошо видны (какой–то исследователь сравнил такое необычное ядро с глазом совы). Некоторые нейроны являются типичными тетраплоидными. Для нейрона характерно наличие специфических структур: нейрофибрилл и хроматофильной субстан-ции. Хроматофильная субстанция (вещество Ниссля или тигроид) на самом деле является мощно развитой гранулярной ЭПС, содержащей многочисленные рибосомы и полисомы (в нервной клетке содержится 20 млн. молекул РНК). Многочисленные рибосомы в телах нейронов непрерывно синтезируют белки (14 тыс. молекул в секунду), с которыми связана обработка информации. Каждые 20 дней белок серого вещества головного мозга обновляется почти полностью. При повреждении аксона, нервном стрессе, психическом расстройстве вещество Ниссля частично или полностью разрушается и ядро сдвигается на периферию.
Хроматофильная субстанция (вещество Ниссля или тигроид) на самом деле является мощно развитой гранулярной ЭПС, содержащей многочисленные рибосомы и полисомы (в нервной клетке содержится 20 млн. молекул РНК). Многочисленные рибосомы в телах нейронов непрерывно синтезируют белки (14 тыс. молекул в секунду), с которыми связана обработка информации. Каждые 20 дней белок серого вещества головного мозга обновляется почти полностью. При повреждении аксона, нервном стрессе, психическом расстройстве вещество Ниссля частично или полностью разрушается и ядро сдвигается на периферию.